Lucid dreaming: Evidence that REM sleep can support unimpaired cognitive function and a methodology for studying the psychophysiology of dreaming
Stephen LaBerge
ABSTRACT: Lucid dreaming provides a test case for theories of dreaming. For example, whether or not "loss of self-reflective awareness" is characteristic of dreaming, it is clearly not necessary to dreaming. Theories of dreaming that do not account for lucidity are incomplete, and theories that do not allow for lucidity are incorrect. The fact that lucid dreamers can remember to perform predetermined actions and signal to the laboratory allows them to mark the time of particular dream events accurately, allowing experiments to establish precise correlations between physiology and subjective reports, and enabling the methodical testing of hypotheses.
Just as dreaming provides a test case for theories of consciousness, lucid dreaming provides a test case for theories of dreaming. Although one is not usually explicitly aware that one is dreaming while in a dream, a remarkable exception sometimes occurs in which one possesses clear cognizance that one is dreaming. During such "lucid" dreams, one can reason clearly, remember the conditions of waking life, and act upon reflection or in accordance with plans decided upon before sleep. These cognitive functions, commonly associated only with waking consciousness, occur while one remains soundly asleep and vividly experiencing a dream world that is often nearly indistinguishable from the "real world" (LaBerge 1985). Theories of dreaming that do not account for lucidity are incomplete, and theories that do not allow for lucidity are incorrect.
Although lucid dreams have been reported since Aristotle, until recently many researchers doubted that the dreaming brain was capable of such a high degree of mental functioning and consciousness. Based on earlier studies showing that some of the eye movements of REM sleep corresponded to the reported direction of the dreamer's gaze (e.g., Roffwarg et al. 1962), we asked subjects to carry out distinctive patterns of voluntary eye movements when they realized they were dreaming. The prearranged eye movement signals appeared on the polygraph records during REM, proving that the subjects had indeed been lucid during uninterrupted REM sleep (LaBerge 1990; LaBerge et al. 1981). Figure 1 shows an example.
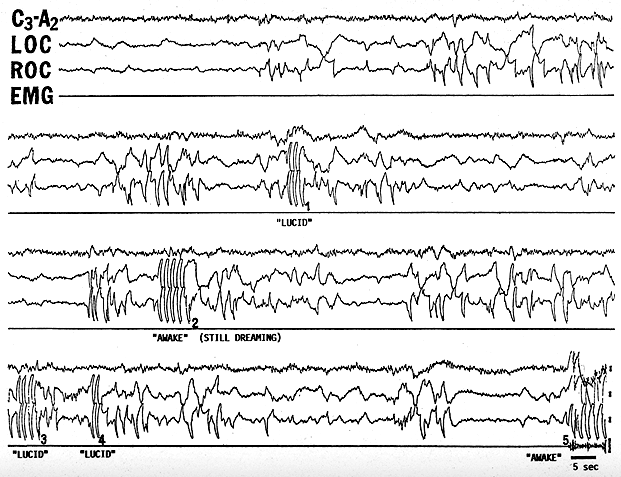
Figure 1. A typical signal-verified lucid dream. Four channels of physiological data (central EEG [C3-A2], left and right eye-movements [LOC and ROC], and chin muscle tone [EMG]) from the last 8 min of a 30 min REM period are shown. Upon awakening the subject reported having made five eye movement signals (labeled 1-5 in figure). The first signal (1, LRLR) marked the onset of lucidity. Skin potential potential artifacts can be observed in the EEG at this point. During the following 90 s the subject "flew about" exploring his dream world until he believed he had awakened, at which point he made the signal for awakening (2, LRLRLRLR). This signal, made in non-lucid REM shows that the precise correspondence between eye movements and gaze is not an artifact of lucidity. After another 90 s, the subject realized he was still dreaming and signaled (3) with three pairs of eye movements. Realizing that this was too many, he correctly signaled with two pairs (4). Finally, upon awakening 100 s later he signaled appropriately (5, LRLRLRLR). [Calibrations are 50 microV and 5 s.]
Our studies of the physiology of lucid dreaming fit within the psychophysiological paradigm of dream research that Hobson has helped establish. Therefore, I naturally agree with Hobson et al. in believing it worthwhile to attempt to relate phenomenological and physiological data across a range of states including waking, NREM, and REM sleep. I also share Hobson's view that REM sleep is unique in many ways; for example, stable lucid dreams appear to be nearly exclusively found in REM. As for the AIM model on which the Hobson et al. article focuses, I regard it as an improvement on the earlier Activation-Synthesis model. The AIM model makes many plausible and interesting connections, but still doesn't do justice to the full range and complexity of the varieties of dreaming consciousness accompanying REM sleep.
One of the problems with AIM is that its three "dimensions" are actually each multidimensional. For example, from which brain area is "Activation" (A) measured? Obviously, A varies as a function of brain location. Hobson et al. admit as much when they propose to locate lucid dreaming in a dissociated AIM space with PFC more activated than it usually is (see Hobson et al.'s Fig. 12). If this is true, then non-lucid dreaming would have to be characterized by a low value of A. Incidentally, there is no evidence to support the idea that lucid dreaming is in any sense a dissociated state (LaBerge 1990). Still, the need for multiple A dimensions seems inescapable.
Similarly, the "Information flow" (I) dimension is more complex than at first appears. Experimental evidence suggests that it is possible for one sense to remain awake, while others fall asleep (LaBerge 1990). A further problem with the I "dimension" is the confounding of sensory input and motor output, as can be seen in several of Hobson et al.'s examples (e.g., compare Figs. 15, 16B, 19).
Finally, "Mode of information processing" (M) attempts to reduce the vast neurochemical complexity of the brain to the global ratio of discharge rates of aminergic to cholinergic neurons. Is that really all there is to say about the neurochemical basis of consciousness? What about regional differences of function? What about the scores of other putative neurotransmitters and neuromodulators?
Perhaps due in part to the over-simplifications necessary to fit these multiple dimensions into an easy-to-visualize three, certain features of dreaming consciousness are misunderstood or exaggerated. For example, Hobson et al. say "self-reflection in dreams is generally found to be absent (Rechtschaffen 1978) or greatly reduced (Bradley et al. 1992) relative to waking." However the two studies cited suffered from weak design and extremely small sample sizes. Neither in fact actually compared frequencies of dreaming reflection to equivalent measures of waking reflection. A study that did make direct comparisons between dreaming and waking (LaBerge, Kahan, & Levitan 1995) found nearly identical frequencies of reflection in dreaming (81%) as in waking (79%), clearly contradicting the characterization of dreams as non-reflective. Replications found similar results (Kahan & LaBerge 1996; Kahan, LaBerge, Levitan, & Zimbardo, 1997). These studies were cited in Hobson's article but otherwise ignored.
Another unsubstantiated claim of Hobson et al. is that "volitional control is greatly attenuated in dreams...." Of course, during non-lucid dreams people rarely attempt to control the course of the dream by magic. The same is true, one hopes, for waking. But likewise, during dreams and waking, one has similar control over one's body and is able to choose, for example to walk in one direction or in another. Such trivial choice is probably as ubiquitous in dreams as waking and, as measured by the question "At any time did you choose between alternative actions after consideration of the options?", 49% of dream samples had voluntary choice, compared to 74% of waking samples (LaBerge et al. 1995). The lower amount of choice in dreams may be an artifact of poorer recall or a real difference, but choice is by no means "greatly attenuated."
While making the above claim, Hobson et al. incorrectly attribute to me the false statement that "the dreamer can only gain lucidity with its concomitant control of dream events for a few seconds (LaBerge 1990)." [sic.] In fact, lucid dreams as verified in the laboratory by eye-movement signalling last up to 50 minutes in length, with the average being about 2 minutes (LaBerge 1990). The relatively low average is partially due to the fact that subjects were carrying out short experiments and wanted to awaken with full recall. At the onset of lucid dreams there is an increased tendency to awaken, probably due to the fact that lucid dreamers are thinking at that point, which withdraws attention from the dream, causing awakening (LaBerge 1985).
The eye-movement signalling methodology mentioned above forms the basis for a powerful approach to dream research: Lucid dreamers can remember pre-sleep instructions to carry out experiments marking the exact time of particular dream events with eye movement signals, allowing precise correlations between the dreamer's subjective reports and recorded physiology, and enabling the methodical testing of hypotheses. We have used this strategy in a series of studies demonstrating a higher degree of isomorphism between dreamed actions and physiological responses than had been found previously using less effective methodologies. For example, we found that time intervals estimated in lucid dreams are very close to actual clock time (see Fig. 2); that dreamed breathing corresponds to actual respiration (Fig. 3); that dreamed movements result in corresponding patterns of muscle twitching (Fig. 4); and that dreamed sexual activity is associated with physiological responses very similar to those that accompany actual sexual activity (see LaBerge 1985, 1990 for details).
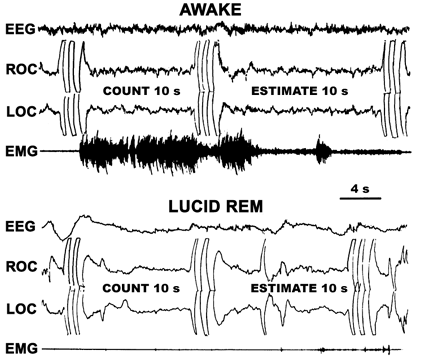
Figure 2. Dream time estimations. We have straightforwardly approached the problem of dream time by asking subjects to estimate ten second intervals (by counting, "one thousand and one, one thousand and two, etc.") during their lucid dreams. Signals marking the beginning and end of the subjective intervals allowed comparison with objective time. In all cases, time estimates during the lucid dreams were very close to the actual time between signals (LaBerge, 1980a, 1985).
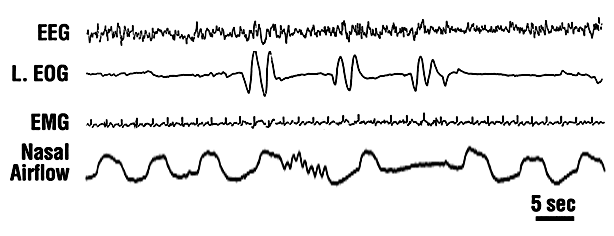
Figure 3. Voluntary control of respiration during lucid dreaming. LaBerge and Dement (1982) recorded three lucid dreamers who were asked to either breathe rapidly or to hold their breath (in their lucid dreams), marking the interval of altered respiration with eye movement signals as shown in the figure. The subjects reported successfully carrying out the agreed-upon tasks a total of nine times, and in every case, a judge was able to correctly predict on the basis of the polygraph recordings which of the two patterns had been executed (binomial test, p < .002).
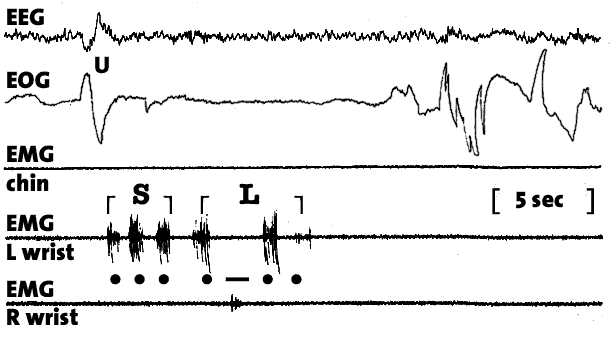
Figure 4. Morse code communication from the lucid dream. Evidence of voluntary control of other muscle groups during REM was found by LaBerge, Nagel, Dement, and Zarcone (1981) while testing a variety of lucidity signals. We observed that a sequence of left and right dream-fist clenches resulted in a corresponding sequence of left and right forearm twitches as measured by EMG. Here the subject sends a Morse code signal with left and right fist clenches corresponding to dots and dashes, respectively. Hence the message translates as "SL" (... .-..), the subject's initials. Note that the amplitude of the twitches bore an unreliable relationship to the subjective intensity of the dreamed action. Because all skeletal muscle groups except those that govern eye-movements and breathing are profoundly inhibited during REM sleep, it is to be expected that most muscular responses to dreamed movements will be feeble. Nonetheless, these responses faithfully reflect the motor patterns of the original dream.
These and related studies show clearly that in REM sleep, dreamed bodily movements generate motor output equivalent at the supraspinal level to the patterns of neuronal activity that would be generated if the corresponding movements were actually executed. Most voluntary muscles are, of course, paralyzed during REM, with the notable exceptions of the ocular and respiratory muscles. Hence, the perfect correspondence between dreamed and actual movements for these two systems (Figs. 1-3), and the attenuated intensity (but preserved spatio-temporal pattern) of movements observed in Figure 4.
These results support the isomorphism hypothesis (Hobson et al.) but contradict Solms's (1995) notion of the "deflection" of motor output away from the usual pathways, and his speculation that it isn't only the musculo-skeletal system that is deactivated during dreams, but "the entire motor system, including its highest psychological components which control goal-directed thought and voluntary action." (Solms 1995, p. 58) I believe Occam's Razor favors the simpler hypothesis that the motor system is working in REM essentially as it is in waking, except for the spinal paralysis; just as the only essential difference between the constructive processes of consciousness in dreaming and waking is the degree of sensory input. See LaBerge (1998) for details.
Oddly, Hobson et al. ignore these data on eye movements while appealing that we keep open the question of relationship between eye movement and dream imagery "until methods more adequate to its investigation are developed." There is no need to wait. Adequate methods have already been developed, as shown above (Figs. 1-3), and in our recent study showing smooth tracking eye movements during dreaming (LaBerge & Zimbardo 2000).
Memory is another area of inquiry upon which lucid dreaming can shed light. Hobson et al. argue that memory during dreaming may be as deficient as it is upon awakening. They give the example of comparing one's memory of a night's dreaming to the memory of a corresponding interval of waking; unless it was a night of drinking being remembered, the dream will yield much less memory. But this is an example comparing episodic memory from waking and dreaming after awakening, and thus is not only unconvincing and vague, but irrelevant. Nobody disagrees that waking memory for dreams is sometimes extremely poor.
In the same vein, Hobson et al. write that it is common for dreams to have scene shifts of which the dreamer takes little note. "If such orientational translocations occurred in waking, memory would immediately note the discontinuity and seek an explanation for it." Note the unquestioned assumption regarding waking consciousness. In fact, recent studies suggest that people are less likely to detect environmental changes than commonly assumed (Mack & Rock 1998). For example, a significant number of normal adults watching a video failed to notice changes when the only actor in a scene transformed into another person across an instantaneous change in camera angle (Levin & Simons 1997).
Likewise, Hobson et al. assert that "there is also strong evidence of deficient memory for prior waking experience in subsequent sleep." However, the evidence offered is always extremely indirect and unconvincing. A direct test requires lucid dreamers to attempt memory tasks while dreaming, as was done in a pilot study (Levitan & LaBerge 1993) showing that about 95% of the subjects could remember in their lucid dreams a key word learned before bed, as well as the time they went to bed, and where they were sleeping. Subjects forgot to do the memory tasks in about 20% of their lucid dreams. That may or may not represent a relative deficit in memory for intentions.
A major methodological difficulty presented by dreaming is poor recall on awakening. The fact that recall for lucid dreams is more complete than for non-lucid dreams (LaBerge 1985) presents another argument in favor of using lucid dreamers as subjects. Not only can they carry out specific experiments in their dreams, but they are also more likely to be able to report them accurately. That our knowledge of the phenomenology of dreaming is severely limited by recall is not always sufficiently appreciated. For example, Hobson et al. repeatedly substitute "dreaming" for "dream recall" (e.g., Sect. 2.3.1). Solms (1997) makes the same mistake, which in my view, is fatal to his argument. So when he writes "of the 111 published cases ... in which focal cerebral lesions caused cessation or near cessation of dreaming...", he is really saying "in which lesions caused cessation of dreaming or dream recall." To think otherwise would be to suppose that the dream is the report.
REFERENCES
Kahan, T.L. & LaBerge, S., (1996). Cognition and metacognition in dreaming and waking: Comparisons of first and third-person ratings. Dreaming, 6, 235-249.
Kahan, T.L., LaBerge, S., Levitan, L., & Zimbardo, P. (1997). Similarities and differences between dreaming and waking: An exploratory study. Consciousness & Cognition, 6, 132-147.
LaBerge, S. (1985). Lucid dreaming. Los Angeles: J. P. Tarcher.
LaBerge, S. (1990). Lucid dreaming: Psychophysiological studies of consciousness during REM sleep. In R.R. Bootsen, J.F. Kihlstrom, & D.L. Schacter (Eds.), Sleep and Cognition. Washington, D.C.: APA Press (pp. 109-126). http://www.lucidity.com/SleepAndCognition.html
LaBerge, S. (1998). Dreaming and Consciousness. In S. Hameroff, A. Kaszniak, & A. Scott (Eds.), Toward a Science of Consciousness II (pp. 494-504). Cambridge, MA: MIT Press.
LaBerge, S., & Dement., W. (1982). Voluntary control of respiration during REM sleep. Sleep Research, 11, 107.
LaBerge, S. & Zimbardo, P.G. (2000). Smooth tracking eye-movements discriminate both dreaming and perception from imagination. Tucson. Paper presented at the Toward a Science of Consciousness Conference, Tucson, April 10-15. http://www.lucidity.com/Tucson2000abs.html
LaBerge, S., Kahan, T. & Levitan, L. (1995). Cognition in dreaming and waking. Sleep Research, 24A, 239.
LaBerge, S., Nagel, L., Dement, W., & Zarcone, V. (1981). Lucid dreaming verified by volitional communication during REM sleep. Perceptual & Motor Skills, 52, 727-732.
Levitan, L., & LaBerge, S. (1993). Dream times and remembrances. NightLight, 5(4), 9-14.
Levin, D.T. & Simons, D.J. (1997). Failure to detect changes to attended objects in motion pictures. Psychonomic Bulletin & Review, 4(4), 501-506.
Mack, A. & Rock, I. (1998). Inattentional blindness. Cambridge, MA: MIT Press.